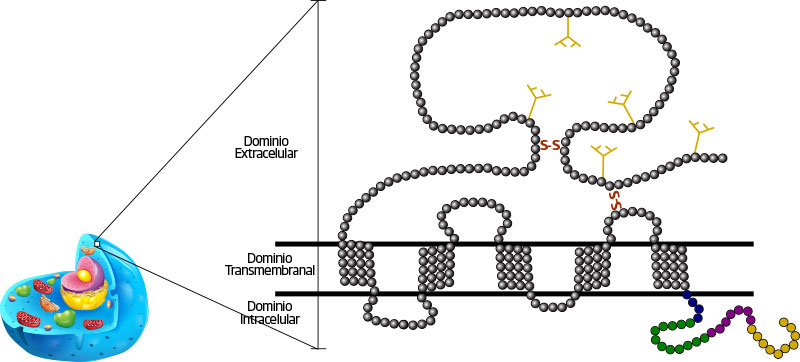
CD47 (figura 1) es una proteína que se encuentra en la membrana de casi todas las células del cuerpo. Funciona como un receptor transmembranal que media de diferentes formas la interacción de las células, comunicando o transduciendoI las señales captadas en el exterior, hacia el interior de la célula. FIGURA 1. Estructura de CD47. Dominio IgV extracelular, con sus cinco sitios de glicosilación (en azul), seguido de los cinco segmentos transmembranales (en gris). Se señalan también los puentes de disulfuro (s-s en rojo) y, por último, el dominio intracelular, donde se encuentra el extremo carboxilo terminal citoplasmático.
CD47 es una proteína de tipo Jano (dios romano con dos caras opuestas), lo cual indica que desempeña dos roles contrarios en diferentes situaciones: En presencia de cáncer, una de sus caras fue descubierta al ver que muchos tipos de células cancerosas expresan grandes cantidades de CD47, lo cual sugiere su complicidad con la enfermedad. Después se comprobó que, de igual manera, estaba implicada en la activación de la MCR en células cancerosas. Dicho de otra manera: CD47 contribuye tanto a la progresión del cáncer como puede inducir su eliminación. ¿Cómo es esto posible? Es lo que trataremos de aclarar en los próximos párrafos, destacando el hecho de que la comprensión de estas dos caras ha contribuido a generar dos potenciales alternativas terapéuticas para desarrollar tratamientos contra el cáncer.

CD47: Un prometedor blanco terapeútico
contra el cáncer
CD47: Un prometedor blanco terapeútico
contra el cáncer
Los diferentes tipos de células de nuestro cuerpo se congregan en poblaciones que mantienen el equilibrio entre la reproducción y la muerte celular. Tanto la proliferación como la muerte son decisiones dinámicas de la célula, activadas por comandos o programas genéticos que permiten mantener un número adecuado de células para que todos los tejidos funcionen con normalidad. Cuando la célula acumula mutaciones que provocan, ya sea un aumento descontrolado de su reproducción y/o la evasión de los programas de muerte celular regulada (MCR) convencionales, este equilibrio se pierde y surge el cáncer.
Inducir la muerte de las células de cáncer —mediante quimioterapias, radioterapias o inmunoterapias— es la estrategia principal para tratar esta enfermedad. La falta de especificidad de los tratamientos para las células cancerosas es un problema grave, por lo que fármacos de nueva generación se dirigen a los componentes específicos de la célula que están involucrados en el desarrollo del cáncer.
Ejemplos de estos componentes son las proteínas —moléculas conformadas por largas cadenas de aminoácidos— que se distribuyen por toda la célula y fuera de ella. Entre sus funciones, están el ser mensajeros o receptores, ya sea en el interior o en el exterior de la célula, y generar señales que promuevan o contrarresten el desarrollo del cáncer. Una proteína particularmente interesante es CD47, ya que desempeña una función clave en el sistema inmune y en la formación de cáncer.
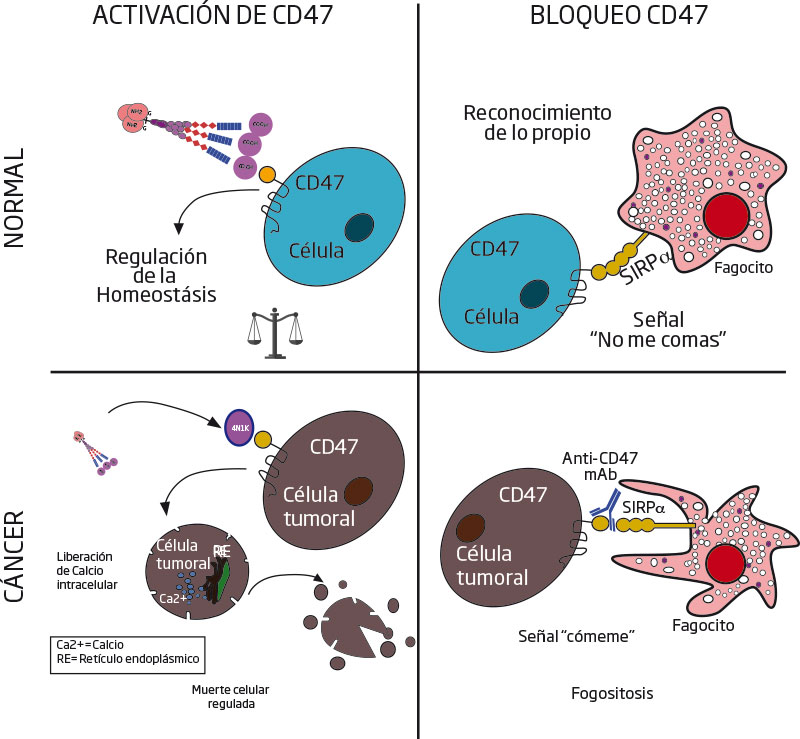
Esto ocurre así por ser el principal receptor de dos proteínas: la trombospondina-1 (TSP1) y la proteína alfa reguladora de señales (SIRP-α).1 La primera, TSP1, es producida y secretada por células del sistema inmune, y se deposita en el espacio denominado matriz extracelular que separa las células unas de otras. La TSP1 se une y activa a CD47 en cualquier célula, mediando la transducción de señales. Por otro lado, la segunda SIRP-α es un receptor transmembranalIII que se expresa sólo en ciertas células del sistema inmune, conocidas como fagocitos profesionales. Estas células devoran a otras, eliminando así bacterias, células infectadas —o potencialmente dañinas, como las cancerosas—. Mediante su interacción con SIRP-α, CD47 funciona como una etiqueta para células propias del cuerpo que claman: “¡no me comas!” a los fagocitos.1, 2, 3, 4 FIGURA 2. CD47 como blanco terapéutico. En células normales, la interacción CD47-TSP1 ayuda a la homeostasis, mientras que la unión CD47- SIRP-α ayuda al reconocimiento de lo propio, evitando la fagocitosis. Por otro lado, en células tumorales, TSP1 y sus péptidos generan MCR, mientras que los anticuerpos monoclonales bloquean la interacción CD47- SIRP-α, estimulando la fagocitosis.
En la pasada década de los noventas, ya se sospechaba la existencia de ambas funciones de CD47; sin embargo, las limitadas tecnologías para la producción de moléculas solubles, además de la constante mejora en la producción de anticuerpos monoclonales (AcMc), favorecieron el desarrollo de la estrategia conocida como “remoción celular programada”.5 El razonamiento es: si CD47 se sobre-expresa en muchos tipos de células de cáncer, evitando así ser reconocida como maligna y removida por los fagocitos, entonces bloquear a CD47 permitiría el reconocimiento de la célula cancerosa y restauraría dicha eliminación.
El bloqueo de CD47 se logró utilizando AcMc, que son proteínas únicas, de gran tamaño (pesan tres veces lo que CD47), producidas por clones específicos de células generadas artificialmente, capaces de reconocer de manera específica una parte de otra proteína —en este caso, la región extracelular de CD47—.
El AcMc dirigido a CD47 más utilizado es B6H12, fue de gran utilidad para validar la hipótesis de que esta estrategia podía ser utilizada para tratar el cáncer. Estudios realizados por el equipo de Weissman y colaboradores usando células de leucemia mielocítica aguda, cáncer de ovario, linfoma no hodgkin, entre otros, mostraron que el bloqueo de CD47 con B6H12 impide su interacción con SIRP-α, y mejora la capacidad de los fagocitos para eliminar células de los diferentes tipos de cáncer estudiados.1
La estrategia basada en este principio está convirtiéndose en una realidad, ya que se generó un AcMc capaz de bloquear la interacción CD47– SIRP-α y permitió la remoción de ciertos tumores sin generar rechazo por parte del sistema inmune en monos. Este nuevo anticuerpo, nombrado Hu5F9-G4, se encuentra ya en las primeras fases de prueba en humanos,3 sin embargo, CD47 no sólo se expresa en células de cáncer, sino prácticamente en cualquier célula. Y, aunque la sobreexpresión de esta proteína en células de cáncer aumenta la probabilidad de que estos AcMc se unan a ellas, también podrían unirse a CD47 en células sanas, como eritrocitos y plaquetas. Esto pudiera provocar efectos secundarios como anemia, trombosis e hipertensión, por lo cual esta estrategia aún se encuentra en estudios clínicos.
Al utilizar B6H12 también se observó que CD47 podía activarse en ciertos escenarios celulares, induciendo MCR. Se descubrió que, de manera fisiológica, dicha acción de CD47 es dirigida por la TSP1,2 haciendo surgir el interés por utilizar pequeñas moléculas llamadas péptidos para generar esta activación.
Los péptidos son porciones pequeñas de proteínas que conservan su capacidad para interactuar con otras. El péptido 4N1K, por ejemplo, conserva la pequeña región de la TSP1 que se une y activa a CD47. Este péptido es el más utilizado en investigación para imitar dicha interacción.
Actividades celulares atribuidas a esta interacción, tales como la adhesión celular en la agregación de plaquetas (el primer paso de la coagulación), la generación de respuestas de la célula ante el estrés, y la activación de la MCR de células cancerosas, han sido propuestas gracias a la utilización del 4N1K.
Recientemente, se creó el péptido PKHB1, conformado por la misma secuencia de aminoácidos que el 4N1K, con pequeñas modificaciones que le confieren mejoras significativas. El PKHB1 es soluble en agua y es más estable en suero que el 4N1K, esto debido a que tiene aminoácidos sintéticos (que no existen naturalmente) en sus extremos, haciendo que no sea cortado por proteínas del suero sanguíneo. A menores concentraciones que su predecesor, este péptido induce MCR selectivamente en células extraídas de la sangre de pacientes con leucemia, pero no en células sanas.4 Esto ocurre, incluso, en las células de pacientes en quienes el tratamiento convencional de primera línea ya no genera respuesta. Además, logró reducir el tamaño de un tumor en ratones sin daño en hígado o riñones.
Otro estudio, además, confirmó que el PKHB1 es también efectivo en células de otros tipos de cáncer,5 sin embargo, las concentraciones de péptido utilizadas en estos ensayos todavía distan de los requerimientos necesarios para el desarrollo de fármacos, las cuales son mucho menores.
En definitiva, utilizar péptidos activadores de CD47 como tratamiento en humanos aún requiere de una ardua labor científica para hacerse realidad. Sin embargo, estos estudios nos hacen ver con optimismo la posibilidad de usar esta estrategia como un futuro tratamiento que podría reducir drásticamente los efectos secundarios de las terapias actuales y funcionar aun en casos de resistencia a los tratamientos de primera línea.

- 4N1K: péptido derivado de la trombospondina 1 que se une a CD47 y ha sido ampliamente utilizado para estudiar la función de CD47, al unirse a la TSP1.
- Anticuerpos monoclonales: anticuerpos idénticos que son producidos por un clon específico de células B inmortalizadas (hibridoma).
- Célula neoplásica: célula con anomalías genéticas o epigenéticas que constituyen una neoplasia o tumor maligno y se dividen a un ritmo más rápido de lo normal.
- Dominio IgV extracelular: región de una proteína transmembranal que se encuentra en el exterior de la célula y tiene similitud con la región variable de las inmunoglobulinas.
- Extremo carboxilo terminal citoplasmático: región final de una proteína transmembranal, cuyo grupo carboxilo (-COOH) se encuentra en el interior del citoplasma.
- Glicosilación: proceso bioquímico en el que se adiciona un glúcido a una molécula.
- Muerte celular regulada: proceso mediante el cual la célula activa su propia maquinaria para autodestruirse y puede ser modulado genética o farmacológicamente.
- SIRP-α: receptor transmembranal presente en fagocitos profesionales que, al interactuar con CD47 en la célula blanco, evita que esta última sea fagocitada.
- Sobre-expresar: presencia de una proteína en la célula en cantidades más altas de lo que ocurre normalmente en células sanas.
- Transducción de señales: transformación de una señal extracelular susceptible de ser detectada por la célula, la cual genera una serie de procesos que desembocan en otra señal o respuesta.
Es Doctora en Fisiología y Fisiopatología por la Universidad Pierre y Marie Curie, y es responsable de la Unidad de Muerte Celular Regulada en Cáncer y el Sistema Inmune, del Laboratorio de Inmunología y Virología, de la Facultad de Ciencias Biológicas, UANL. Su investigación se ha enfocado en el estudio y la activación de la muerte celular regulada como estrategia para combatir las células cancerosas. Es miembro del SNI, nivel 1.
C. e.: ana.martinezto@uanl.edu.mx
Es Maestra en Ciencias con orientación en Inmunobiología y becaria Nacional PNPC Conacyt del Doctorado en Ciencias con orientación en Inmunobiología, de la Facultad de Ciencias Biológicas, UANL.
Es Maestro en Ciencias con orientación en Inmunobiología, y aspirante al Doctorado en Ciencias con orientación en Inmunobiología, de la Facultad de Ciencias Biológicas, UANL.
Es Doctora en Ciencias con especialidad en Inmunología por el Instituto Politécnico Nacional. Es jefe del Laboratorio de Inmunología y Virología, y coordinadora del Posgrado en Inmunobiología de la Facultad de Ciencias Biológicas, UANL. Tiene amplia experiencia en Investigación Biomédica enfocada a la inmunología y al cáncer. Cuenta con patentes y artículos relacionados con nuevas terapias enfocadas en estimular el sistema inmune y en activar vías de muerte celular con el fin de eliminar células tumorales. Es miembro del SNI (III).